Based on the article: Immunization of Experimental Dogs With Salivary Proteins From Lutzomyia longipalpis, Using DNA and Recombinant Canarypox Virus Induces Immune Responses Consistent With Protection Against Leishmania infantum, Abbehusen et al. 2018
Severe and sometimes lethal diseases can be transmitted from animals to humans, creating a high risk factor for human infection. One such disease, visceral leishmaniasis (VL), is caused by infection with Leishmania infantum [1]. Leishmania infantum is the causative agent of VL that is transmitted through the bite of infected sand flies, Lutzomyia longipalpis, during blood feeding. Domestic dogs are commonly affected by the bite of these sand flies in the Mediterranean Basin, South America, and parts of Asia [2]. This transmission is particularly dangerous because it can be transmitted from domestic dogs to human owners, resulting in severe disease and potential death. Therefore, in order to combat the spread of VL disease, the authors developed an immunization for the dogs using salivary proteins from the sand flies that are known to carry the causative agent of the disease, Leishmania infantum.
Immunizations are essential for creating immunity or resistance to a particular disease. In a previous study, the authors of this paper showed that the use of DNA plasmids encoding a specific salivary protein from the sand fly that transmits the disease, Lutzomyia longipalpis, induced a strong immune response that provided protection against the Leishmania infantum bacteria in hamsters [3]. In other words, a salivary protein was taken from the parasite that carries the disease to immunize hamsters against the disease. The main concept behind this process was that the hamsters were initially exposed to salivary proteins from the sand fly that caused the immune system to mount a response so that when the hamster was exposed to the harmful L. infantum carried by the sand fly, the immune system already had the necessary antibodies to fight it off.
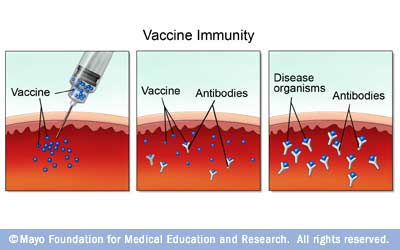
Figure 1. Schematic diagram of how vaccines work. A vaccine is injected and bound by antibodies so that when the pathogen invades the host, there are already antibodies present to fight it off.
Based on the success of the L. longipalpis immunization in hamsters against L. infantum, the authors developed a similar experimental procedure to test on dogs to induce immunity against the VL disease. Two L. longipalpis salivary proteins, LJM17 and LJL143, were identified and used in an immunization strategy to see if a strong enough immune response would be generated to protect the dogs from L. infantum infection. In order to test the effect of these salivary proteins on inducing immunity to L. infantum infection in dogs, the authors gave a single dose of plasmids encoding either the LJM17 or the LJL143 salivary protein, followed by two additional booster doses of recombinant Canarypoxviruses vector expressing one of the two salivary protein genes. These vectors have proven to be safe for vaccination use [4]. Then, the dogs were experimentally infected with L. infantum in the presence of L. longipalpis saliva in order to gage the success of the vaccination in protecting the dogs against L. infantum infection.
Beagle dogs were split into two treatment groups for either the LJM17 or the LJL143 salivary protein in addition to a control group treated with saline solution. The dogs were first exposed only to the immunization without L. infantum. Then, in another series of experiments, the dogs were infected with L. infantum in the presence of the salivary proteins. The results of exposure to the salivary proteins alone demonstrated that dogs immunized with LJM17 and LJL143 proteins presented a mixed immune response 15 days after the last immunization. This means that the immune response included components of both a humoral and cell mediated immune response. A humoral immune response involves the secretion of antibodies, while a cell mediated immune response involves the activation of phagocytes, cytotoxic T cells, and the release of various cytokines [5, 6]. The experimental groups that were immunized with the two salivary proteins showed production of specific antibodies against the two proteins when compared to the control group, demonstrating the ability of the proteins to provoke an immune response in the dogs (Figure 2A and 2B). The production of IgG1 and IgG2 antibodies indicated the presence of a humoral response against the salivary proteins (Figure 2C). IgG antibodies are the most common type of antibody found in blood circulation and are created and released by plasma B cells. Therefore, the presence of IgG antibodies indicates the occurrence of a humoral response, in which antibodies are produced to destroy extracellular microorganisms and prevent the spread of infection.

Figure 2. Analysis of humoral and cellular immune response in immunized and control dogs. Sera collected 15 days after the last immunization were used to detect total IgG (A) anti-LJM17 (B) anti-LJL143 and (C) specific IgG1/IgG2 subclasses.
In addition to antibody production, the authors also measured cytokine production 15 days after the last immunization in the dogs. The authors found that immunized dogs exhibited higher IFN-γ production when compared to controls (Figure 3). This finding is significant because IFN-γ is an important pro-inflammatory Th1 cytokine that is produced during a cell-mediated immune response [7]. IFN-γ differentiates T cells into Th1 cells, which induce a pro-inflammatory immune response. Therefore, the production of IFN-γ in immunized dogs indicates that the immunization successfully resulted in a Th1 immune response in the dogs. The production of antibodies and inflammatory cytokines demonstrates that dogs immunized with salivary proteins presented a mixed immune response with components of both a humoral and cell mediated response.

Figure 3. Analysis of humoral and cellular immune response in immunized and control dogs. Sera collected 15 days after the last immunization were used to detect total IFN-γ production in immunized and control dog sera.
After examining the immune responses to the immunizations, the authors evaluated the immune response induced by infection with L. infantum in the presence of the salivary proteins to see if the proteins were protective against L. infantum. The results demonstrated that at 2 months after infection, anti-L. infantum antibodies had already been detected in all dogs and that the groups that received the LJM17 immunization presented significantly higher levels of IgG in comparison to the control group. The authors also found that the dogs immunized with LJM17 produced significantly higher amounts of IFN-γ when infected with L. infantum, indicating a stronger immune response than the dogs that were not immunized with LJM17.

Figure 4. Detection of IFN-γ in sera from immunized dogs and controls challenged with L. infantum.
In order to evaluate whether the presence of the salivary proteins were protective against the L. infantum infection, the authors assessed parasite load in each experimental and control group of dog sera and found that parasites were only detected in the skin and spleen at 2 months after infection in both immunized and control groups. This lack of difference between immunized and control groups demonstrates that even though the dogs were effectively infected, the authors were not able to confirm any preventative action with regards to the immunization protocol. Although the immunizations clearly induced immune responses in the dogs treated with LJM17, the authors could not confidently report whether the immunizations were effective in protecting against L. infantum because parasites were present in both the experimental and control groups 2 months after infection and they were not present at all at later time points in all groups. Nonetheless, the authors still noticed differences in cytokine production between the experimental groups and control group when infected with L. infantum.
The authors found that immunized dogs expressed additional cytokines following infection, even though it was unclear if the induced immune response was able to protect the dog against the infection. They looked at the cytokines produced directly after the immunization, 2 months after the immunization, and 4 months after the immunization in dogs immunized with LJM17 and LJL143 and compared the results to that of the control group. Overall, the results demonstrated that the dogs immunized with LJM17 presented a pattern of pro-inflammatory responses that persisted until 4 months after infection, as indicated by the production of pro-inflammatory cytokines. In contrast, the results demonstrate that dogs immunized with LJL143 presented an immediate inflammatory response after the immunization that was less intense than that of LJM17-immunized dogs. The dogs given the control saline treatment did not present any increases in inflammatory cytokines. Therefore, even though the parasites were detected in both the immunized and control groups 2 months after the immunization, there was sufficient evidence for the occurrence of immune responses in immunized dogs while there was no immune response in the control group of dogs infected with L. infantum.
While a previous study by Collin et al. demonstrated that a more complex immunization strategy with L. longipalpis salivary proteins LJM17 and LJL143 resulted in systemic and local Th1 cell mediated immunity that interfered with parasite survival in hamsters, this study does not explicitly show the protective effects of LJM17 and LJL143 immunizations against L. infantum infection in dogs [8]. This study conducted by Abbehusen et al. differed because they used dogs instead of hamsters, and a simplified vaccination schedule as opposed to a more complex method used by Collin et al. Although all experimental and control groups were sufficiently infected, the parasites were no longer present in any of the dogs 2 months after immunization. Therefore, it is unclear as to whether the immune responses induced by the immunizations resulted in protection against the infection. Despite this shortcoming of the experiment, the results are important because the immunization strategy elicited a strong humoral and cell mediated immune response. Dogs immunized with LJM17 and LJL143 showed elevated production of IgG antibodies, IFN-γ, and other cytokines, which are similar to the results obtained by Collin et al. Furthermore, when immunized in the presence of the L. infantum infection, the dog sera demonstrated an increase in antibodies against L. infantum. Therefore, the results corroborate previous evidence that these salivary proteins elicit a strong immune response that could potentially be beneficial for fighting off infection. More specifically, the results indicate that the LJM17 immunization induced an immune response with high levels of IFN-γ, which is consistent with known protective immune mechanisms against L. infantum. Therefore, even though the results of this study did not prove the protective effect of LJM17 immunization directly, the immune response it induced suggests that it would serve a protective function against L. infantum.
In future studies, the authors should induce a higher infection pressure to ensure that the dogs are infected enough to develop the VL disease. This would allow for more adequate experimental settings to assess whether the salivary proteins can not only induce an immune response but also limit the development of the disease and produce evidence of a protective effect specifically against L. infantum. Furthermore, the authors should use a larger sample size in future experiments, as only 30 dogs total with 10 in each group were used for the study. A larger sample size and a stronger infection to make sure the VL disease develops would be helpful for a future study on the use of this heterologous prime-boost vaccination strategy with L. longipalpis salivary proteins. Future studies will be necessary to further investigate the potential role of LJM17 as a vaccine component to protect against disease.
Works Cited:
[1] "Visceral leishmaniasis." World Health Organization, https://www.who.int/leishmaniasis/visceral_leishmaniasis/en/.
[2] Abbehusen, Melissa Moura Costa et al. "Immunization of Experimental Dogs With Salivary Proteins From Lutzomyia longipalpis, Using DNA and Recombinant Canarypox Virus Induces Immune Responses Consistent With Protection Against Leishmania infantum." Frontiers in Immunology (2018). https://www.frontiersin.org/articles/10.3389/fimmu.2018.02558/full.
[3] "Bacterial DNA - the role of plasmids." Science Learning Hub, 25 March 2014. https://www.sciencelearn.org.nz/resources/1900-bacterial-dna-the-role-of-plasmids.
[4] Plotkin, SA et al. "The safety and use of canarypox vectored vaccines." Dev. Biol. Stand. (1995). https://www.ncbi.nlm.nih.gov/pubmed/7796950.
[5] Janeway, CA Jr et al. "The Humoral Immune Response." Immunobiology: The Immune System in Health and Disease. (2001) https://www.ncbi.nlm.nih.gov/books/NBK10752/.
[6] Janeway, CA Jr et al. "T Cell-Mediated Immunity". Immunobiology: The Immune System in Health and Disease. (2001)https://www.ncbi.nlm.nih.gov/books/NBK10762/.
[7] "Interferon Gamma." ScienceDirect, 2018. https://www.sciencedirect.com/topics/immunology-and-microbiology/interferon-gamma.
[8] Collin, N et al. "Sand fly salivary proteins induce strong cellular immunity in a natural reservoir of visceral leishmaniasis with adverse consequences for Leishmania." PLoS Pathog. (2009). https://www.ncbi.nlm.nih.gov/pubmed/19461875.
No comments:
Post a Comment