Review based on the paper: Pilling, D., Chinea, L. E., Consalvo, K. M., & Gomer, R. H. (2018). Different isoforms of the neuronal guidance molecule Slit2 directly cause chemoattraction or chemorepulsion of human neutrophils. The Journal of Immunology
Immune cells like neutrophils are spread out all over our body. They circulate in blood like other white blood cells. As part of their function in innate immunity, however, they need to arrive at specific locations of injury, infection, or inflammation. To do so, they use a process called chemotaxis - movement in response to a chemical stimulus. Positive chemotaxis is the movement towards the higher concentration of the chemical (chemoattractant). Negative chemotaxis is the movement away from the higher concentration of the chemical (chemorepellent). Therefore, if the cells at the site of inflammation start secreting chemoattractants, neutrophils will start moving towards higher concentration and it will eventually lead them to the site of inflammation. Chemotaxis is not only used by immune cells. Especially during the development, many tissues like the nervous system use chemorepellents and chemoattractants to control the development in an organized manner. A protein called Slit was originally identified as a neuronal chemorepellent, for example.1 Slit proteins are sensed by Robo receptors and prevent neurons from crossing through the midline of the central nervous system.2 Interestingly, immune cells such as neutrophils also express Robo receptors.3 With this knowledge, a group of researchers attempted to study how Slit is involved in neutrophil chemotaxis and they found that different isoforms of Slit2 (one of the three homologs of Slit in humans) can cause either chemoattraction or chemorepulsion.1 Let’s dive in!
Mammals have three slit homologs: Slit1, Slit2, Slit3.4 Slit2 in mammals is cleaved into N-terminal and C-terminal fragments.4 Depending on how it’s cleaved, the N-terminal fragment is either 110 kDa (Slit2-S) in size or 140 kDa (Slit2-N). Intriguingly, altered cleavage patterns of Slit2 were found in cancer, inflammation, fibrosis, and other immunologically active and relevant sites1 which once again made researchers question whether the Slit2 protein is immunologically relevant in chemotaxis. To test this, researchers designed Slit2 gradient chambers, placed neutrophils onto the gradient, and observed their motion. Figure 1 (B,C) shows mapped and centered trajectories of each neutrophil placed on a Slit2 concentration gradient for different isoforms. Concentration ranges from 0 to 500 ng/ml with the highest concentration on the left edge of the chamber. Each neutrophil starts the movement from the center, and then moves along the line and ends up on a dark spot in 40 mins. In the control experiment with no Slit2 gradient, neutrophils barely move from the starting position (Fig. 1A), whereas neutrophils on the Slit2-S and Slit2-N gradient move away from the starting position (Fig. 1B,C).1 Figure 1 (E,F) presents quantified data of the average direction of neutrophils moving on different concentration gradients of different isoforms of Slit2. Positive FMI indicates the movement away from the Slit2, and negative FMI indicates the movement towards Slit2. Based on their data, Slit2-S acts as chemorepellent within the concentration range of 0-500 ng/ml and so does Slit2-N within the concentration range of 0-5000 ng/ml.1 Slit2-N, however, can act as a chemoattractant when the concentration ranges from 0 to 500 ng/ml (Fig. 1F).1
 |
| Figure 1 |
I personally find the data not as conceiving as I’d like it to be. First of all, even though the FMI of the Slit2-S is comparable to the FMI of other chemorepellents for neutrophils, even just looking at the mapped trajectories one can tell that many of the neutrophils did not move away from the high concentration of the Slit2-S at all. As researchers claim, 30% of all the cells actually moved towards the source of Slit2-S1 which seems a little bit too high of a number for me to support the claim that the Slit2-S is a chemorepellent. Second, if we look at the Fig.1 E panel, we can see that once a chemoattractant fMLP is added to the chamber, neutrophils start moving towards the source of the molecules. This in itself is not bad. It seems like chemoattractant dominates over the effects of Slit-2S chemorepulsion. However, what caught my attention is that cells move closer to the source in the presence of 0-500 ng/ml Slit2-S gradient compared to the 0-5000 ng/ml or 0-50 ng/ml Slit2-S gradients which seems counterintuitive. Chemorepellent force of Slit2-S is the strongest in the 0-500 ng/ml gradient.1 Unfortunately, authors do not really look into this to provide possible explanations. Is there a specific interaction between Slit2-S and fMLP that yield these results? Maybe, but unlikely. Are the experiments conducted poorly that give unreliable results? Maybe!
As for Slit2-N, researchers concluded that in the 0-5000 ng/ml chamber the amount of the cells that moved towards the source was significantly high (38%).1 Even though average FMI indicates that Slit2-N acted as a chemorepellent within this particular concentration range, they decided it is a chemoattractant since in the 0-500 ng/ml chamber average FMI was negative and only 17% of the cells moved away from the source.1 Nevertheless, authors speculate that gradient of Slit2-N that starts at a high concentration might attract neutrophils to tissues but create an exclusion zone near the source of the Slit2-N.1 However, they do not provide any explanation why and how Slit2-N is acting in the opposite way at higher concentrations.
Even though data from Figure 1 leaves us with some lingering questions, Figure 2 does strengthen the findings they present. Both neurons and neutrophils use Robo1 as a receptor to sense Slit2.3,4 Neutrophils preincubated with anti-Robo1 antibodies did not respond to either Slit2-N or Slit2-S gradients (Fig.2 A).1
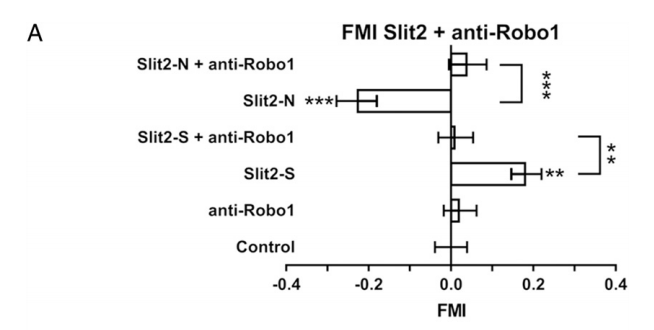 |
| Figure 2 |
Further experiments that the researchers perform show that Slit2-S and Slit2-N have different effects on neutrophils. First, they show that cytoskeleton is affected differently since they induce different polarities of actin polarization.1 Then they show that Slit2 isoforms affect adhesion and cell shape differently. More neutrophils treated with Slit2-N seem to adhere to the coated plate than those treated with Slit2-S or with nothing at all.1 Slit2-N treatment also increases the length and the area of the cells, whereas Slit2-S seems to have no effect on the cell shape.1 In the following experiments, researchers show that different signaling molecules are involved in propagating either chemorepulsion or chemoattraction with Slit2-S or Slit2-N respectively.1 Overall, this led the researchers to believe that the two Slit2 fragments activate different signal transduction pathways despite using the same receptor.1
Overall, despite my critiques of data presented in figure 1, I think this paper manages to present the complexity of the topic well and shows that there’s still a lot of work to be done to understand the roles of Slit2 in the chemotaxis of neutrophils. I think the researchers fail to provide enough evidence to show the importance of their research. For better context, I wish they had provided physiological concentrations of Slit2 fragments just so that we know that they are comparable to the concentrations used in the experiments. Furthermore, at the beginning of the paper authors mentioned that different fragmentation patterns were found by other researchers in different conditions. I wish they had provided an analysis of these patterns. Is more Slit2-N produced at places where neutrophil is homed? Is more Slit2-S produced at places where neutrophil activity is not needed? This analysis might have helped them confirm or reject their findings. This research also leads us to different questions. How is Slit2 cleavage mediated/regulated? Are neuronal and neutrophil chemotaxis functionally related? How are different signals elicited from Robo1? Once more experiments will be done addressing this question, we will have a better, big-picture understanding of the roles of Slit2 proteins in neutrophil activity.
P.S. The title references the psychological thriller Split which is a movie about a person with 23 different personalities :)
References
- Pilling, D., Chinea, L. E., Consalvo, K. M., & Gomer, R. H. (2018). Different isoforms of the neuronal guidance molecule Slit2 directly cause chemoattraction or chemorepulsion of human neutrophils. The Journal of Immunology, , ji1800681. doi:10.4049/jimmunol.1800681
- Zhang, C., Gao, J., Zhang, H., Sun, L., & Peng, G. (2012). Robo2-slit and dcc-Netrin1 coordinate neuron axonal pathfinding within the embryonic axon tracts. Journal of Neuroscience, 32(36), 12589-12602.
- Tole, S., Mukovozov, I. M., Huang, Y., Marco A. O. Magalhaes, Yan, M., Crow, M. R., . . . Robinson, L. A. (2009). The axonal repellent, Slit2, inhibits directional migration of circulating neutrophils. Journal of Leukocyte Biology, 86(6), 1403-1415.
- Blockus, H., & Chedotal, A. (2016). Slit-robo signaling. Development, 143(17), 3037-3044.
No comments:
Post a Comment