As a result of decades of research on HIV virus, we know a lot about its structure. We also know the identity of the main molecular players in host cell invasion. This knowledge has been used to design different therapies like specific antibody treatments, though they have not been the most effective.2 In order to come up with better therapies, we need to know exactly how these molecular players interact that enables HIV molecule to infect us by entering the cells. Shaik et al.1 try to uncover the molecular mechanisms of HIV infection by looking at membrane protein interactions through cryo-electron microscopy.
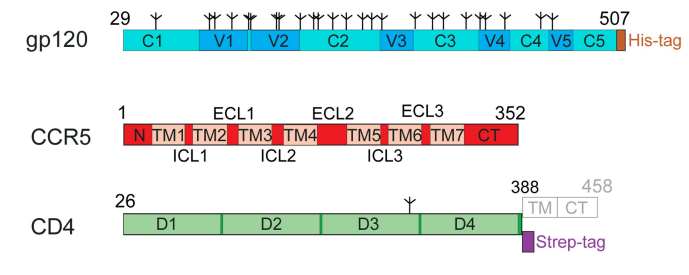
HIV is an enveloped virus, meaning it has a lipid bilayer around it. Major membrane protein for HIV is a glycoprotein gp160 which is cleaved into trimeric gp120 (exposed domain) and gp41 (transmembrane domain).3 Three copies of each fragment constitute the viral spike also called Env. It's been previously believed that interaction of gp120 with the primary receptor CD4 on a host cell and a coreceptor (for example, CCR5 or CXCR4) triggers a conformational change in gp120 followed by the dissociation of gp120.3 This supposedly exposes gp41 which then plays a crucial role in invasion by promoting membrane fusion.3 We now have a good understanding of the molecular dynamics of interaction between gp120 and CD44 but very little is known about the molecular and structural basis of the interaction between Env and a coreceptor.1 To gain a better understanding of this interaction, Shaik et al. determined the structure of CCR5 in complex with an HIV-1 gp120 and a four-domain soluble CD4 by a high-resolution cryo-electron microscopy.1 To obtain such a complex, they treated the cells that overexpress human CCR5 with gp120 in the presence of soluble CD4.1 Then they isolated and purified the CD4-gp120-CCR5 complexes from the cells and acquired cryo-EM images after freezing them.1 (for more information on isolation, purification, and positive controls, read the section CCR5 complex purification and structure determination in the paper).
 |
| Figure 1 |
Before we look at their images and the interactions these researchers have identified let’s point out some of the domains that have previously been found to be important. In gp120, V3 loop seems to be involved in the interaction with CCR5 and CXC4.5 Mutagenesis experiments have also shown that N-terminal extracellular tail is important for binding in CCR5.6 Not all 4 domains of CD4 are involved in the interaction with gp120 but researchers decided to leave all 4 extracellular domains for maximum accuracy in results, however they did not include transmembrane and intracellular domains since they are highly unlikely to be involved in the interactions.1
Based on the images researchers obtained, they identified two main interaction zones between gp120 and CCR5.1 One, the V3 loop of gp120 is inserted into a binding pocket called CRS2 of CCR5 (Fig 2).1 Two, N-terminus of CCR5 interacts with the bridging sheet of gp120 (Fig 2).1
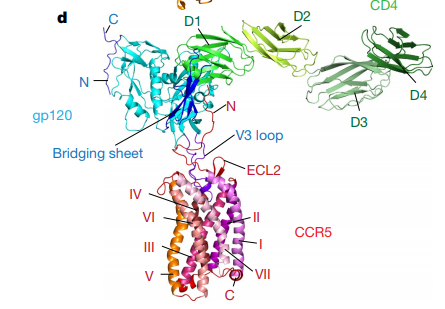 |
| Figure 2: Structure of the CD4-gp120-CCR5 complex. |
1. The V3 loop of gp120 and CRS2 of CCR5.
Upon interaction between Env and its coreceptor, the V3 loop of gp120 inserts into the binding pocket CRS2 which is surrounded by 7 transmembrane segments. V3 makes contact with all the helices of the 7 transmembrane domains of CCR51. Because V3 makes extensive contacts with the coreceptor, authors believe this interaction contributes to the high affinity of HIV-1 Env for CCR5 and CXC4.1 Researchers decided to look at the CRS2 of CCR5 upon binding with different ligands.
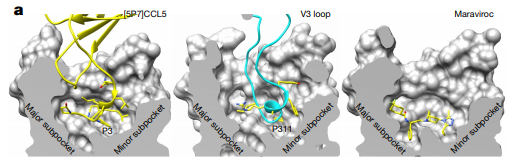 |
| Figure 3: Interaction of CCL5, gp120, and maraviroc with CCR5 in CRS2 binding pocket. |
One of the ligands in figure 3 is maraviroc. Maraviroc is a drug that has previously been believed to inhibit HIV-1 infection by stabilizing the conformation of CCR5 that gp120 cannot recognize.7 It seems like that hypothesis is not right since the conformation of the pocket mostly stayed unchanged (Fig. 3).1 In fact, the structure of the binding site CRS2 differs only minimally when bound by different ligands, suggesting that this part of the CCR5 is pretty rigid.1 On the other hand, Shaik et al. noticed that the binding site of Maraviroc and the space the drug occupies in the pocket partially overlaps with the binding site of the V3 loop tip.1 Therefore, it is very likely that Maraviroc blocks gp120 binding by direct competition.1
2. The gp120 Bridging sheet and CCR5 N terminus.
The second major interaction point between gp120 and its coreceptor is formed between the extracellular N-terminus of CCR5 and the bridging sheet of gp120. Usually, N-terminal tail is floppy and adopts different structures upon binding with other CCR5 ligands. In the CD4-gp120-CCR5 complex, however, N-terminal segment adopts an extended structure with several sharp turns and interacts with the surface of the bridging sheet, a structure of gp120 which is accessible only after CD4 binding.1
The fact that the amino acids on the two proteins interact only on a handful of locations explains why HIV viruses can easily mutate their genome to change the identity of the coreceptor it interacts with.1 Besides identifying and characterizing two major faces of interaction, there are two important findings that the research revealed. First, CD4 lies almost perpendicular to CCR5 (Fig 2).1 Researchers think this orientation of interaction could possibly induce a bend in the membrane (Fig. 5).1 Second, it seems like there is no obvious allosteric change in Env induced by coreceptor binding revealing gp41 as previously it was thought to be.1 A comparison between CD4-bound and CCR5- and CD4-bound gp120 does not reveal any major differences in gp120 structure (Fig.4)(the minor differences in the structural change and their insignificance to gp41 dissociation is discussed further in detail in the paper in sections Differences between CD4- and CCR5-bound pg120 and Model for how CCR5 functions as an HIV-1 coreceptor).1 Hence, Shaik et al. suggest that conformational change that induces gp41 dissociation is followed by gp120-CD4 binding alone.1
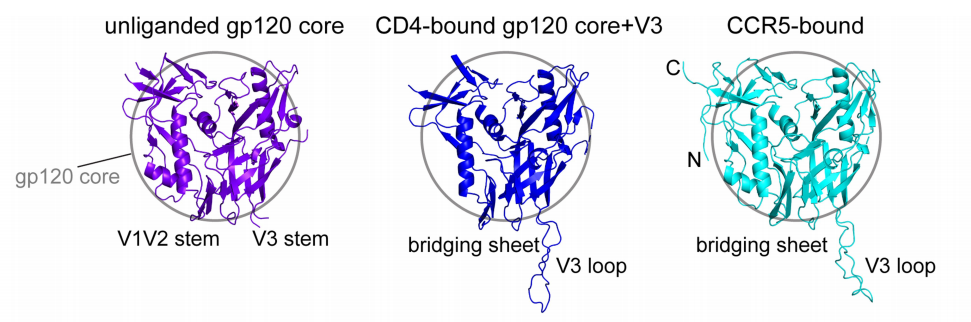 |
| Figure 4: Structure of gp120 when unliganded, CD4-bound, or CD4- and CCR5-bound |
This finding left the researchers wondering how does then CCR5 function as an essential coreceptor. If CCR5-binding is not a molecular mechanism that induced gp41 dissociation from gp120, then why is gp120-coreceptor interaction needed for effective HIV-1 infection? The researchers provide three reasons as to why they think coreceptor is important:
1. The gp120-CCR5 binding brings the fusing site of gp41 as close as 70Å away from the host cell membrane compared to 160Å when Env interacts with CD4 alone.1 The closer distance might be critical to the fusion process. Therefore, coreceptor is important for increasing proximity between the membrane and gp41.1
2. Even though it seems more likely that CD4 induces the necessary conformational change in gp120, apparently, gp120-CD4 association is very weak and, therefore, rapidly reversible.8 Unless it is stabilized, gp41 will not be fully dissociated from gp120.1,8 Coreceptor is thus necessary to stabilize the CD4-induced conformational change, which in itself is sufficient for triggering and promoting fusion.1
3. Based on what we know about other viruses, many Env proteins might be necessary for fusion pore to be formed.9 Coreceptor stabilization might be crucial to keep the Env-CD4 interaction for long enough to give it enough time to recruit other Env-CD4 complexes to the site to form the fusion pore.1
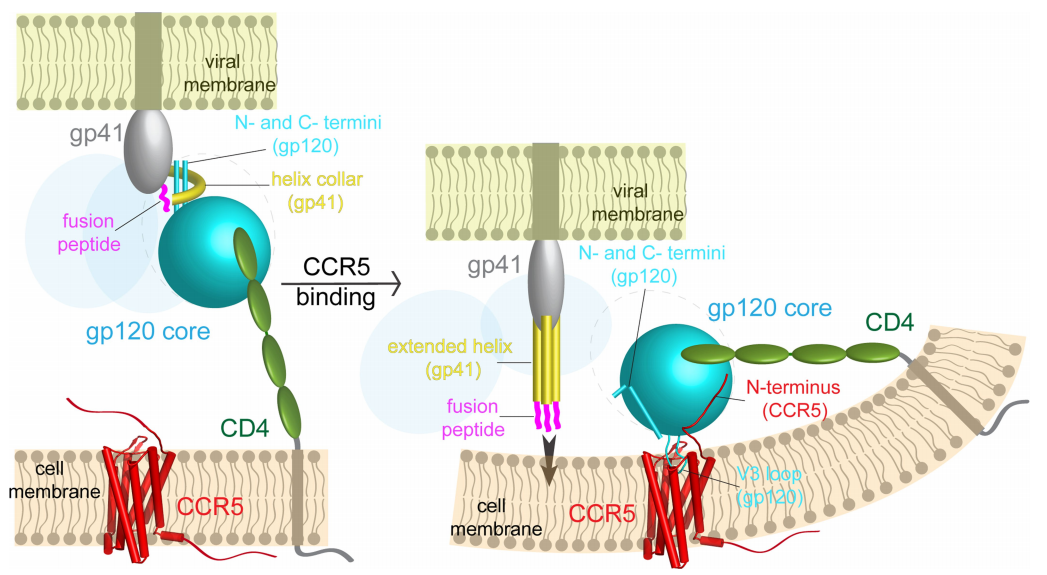 |
| Figure 5: Visual representation of the proposed molecular mechanism |
Overall this research reveals some of the most important details of the molecular mechanism of HIV-1 infection. Future research should concentrate on how this information can be used to design new therapies for infected individuals or how to prevent infection in healthy individuals, to begin with. Now that we know exactly how Maraviroc drug functions, we can also design more effective drugs by intentionally attaching bulkier groups to Maraviroc molecule so that its binding to CRS2 is not impaired but the space it takes up in the pocket increases.1 I, personally, am intrigued by the idea of bending membrane. First of all, I am skeptical that the membrane actually bends that much. We should not reject the possibility that if all the proteins were bound to the membrane (CD4 was not in this experiment), the interactions might have been slightly different or at different angles. Maybe it is not the membrane that bends, but gp120 itself. It is possible that the stress applied by CCR5 and CD4 cause conformational change in gp120 as membrane keeps two transmembrane proteins parallel to each other. If membrane does bend though, what role does this bend play in viral fusion? I am hoping the researchers in this field will try to find answers to these questions as the knowledge they obtain will be directly applied to the drug and therapy design for the infected individuals.
Figures are taken from the original paper.
1.Shaik, M. M., Peng, H., Lu, J., Rits-Volloch, S., Xu, C., Liao, M., & Chen, B. (2018). Structural basis of coreceptor recognition by HIV-1 envelope spike. Nature, doi:10.1038/s41586-018-0804-9
2. Margolis, D. M., Koup, R. A., & Ferrari, G. (2017). HIV antibodies for treatment of HIV infection. Immunological Reviews, 275(1), 313-323.
3. Harrison, S. C. (2008; 2015). Viral membrane fusion. Nature Structural & Molecular Biology, 15(7), 690-698.
4. Wang, H., Cohen, A., Galimidi, R., Gristick, H., Jensen, G., & Bjorkman, P. (2016). Cryo-EM structure of a CD4-bound open HIV-1 envelope trimer reveals structural rearrangements of the gp120 V1V2 loop. Proceedings of the National Academy of Sciences of the United States of America, 113(46), E7151-E7158.
5. Rizzuto, C., Wyatt, R., Hernandez-Ramos, N., Sun, Y., Kwong, P., Hendrickson, W., & Sodroski, J. (1998). A conserved HIV gp120 glycoprotein structure involved in chemokine receptor binding. Science, 280(5371), 1949-1953.
6. Doranz, B. J., Lu, Z. H., Rucker, J., Zhang, T. Y., Sharron, M., Cen, Y. H., . . . Peiper, S. C. (1997). Two distinct CCR5 domains can mediate coreceptor usage by human immunodeficiency virus type 1. Journal of Virology, 71(9), 6305-6314.
7. Moore, J., & Kuritzkes, D. (2009). A piece de resistance: How HIV-1 escapes small molecule CCR5 inhibitors. Current Opinion in Hiv and Aids, 4(2), 118-124.
8. Dobrowsky, T. M., Zhou, Y., Sun, S. X., Siliciano, R. F., & Wirtz, D. (2008). Monitoring early fusion dynamics of human immunodeficiency virus type 1 at single-molecule resolution. Journal of Virology, 82(14), 7022-7033.
9. Floyd, D. L., Ragains, J. R., Skehel, J. J., Harrison, S. C., & Oijen, A. M. v. (2008). Single-particle kinetics of influenza virus membrane fusion. Proceedings of the National Academy of Sciences of the United States of America, 105(40), 15382-15387.
Sands Casino | SA, TENNIS, Canada
ReplyDeleteSands 메리트 카지노 쿠폰 Casino is a casino in TENNIS, Canada, owned and operated by Sands Corp. It 샌즈카지노 is open 인카지노 daily 24 hours from Main Street.