Based on: Secreted IgD amplifies humoral T helper 2 cell responses by binding basophils via Galectin-9 and CD44
Principal Investigator: Meimei Shan, MBBS, PhD
Date of publication: October 2, 2018
Understanding the mechanisms involved in the adaptive immune response is imperative to developing new research techniques and potential cures for diseases and pathogens. This recent study published in Immunology in early October of 2018 outlines the role that IgD plays in T helper 2 (Th2) cell activation and the proliferation of basophils. While the effector functions of immunoglobulin M (IgM), IgG, IgA, and IgE after release by B cells are well documented, the role of IgD has remained unclear.
T Helper 2 cells
T helper cells are extremely important in the adaptive immune response, in that they not only help to activate B cells to secrete antibodies and macrophages to destroy ingested microbes by phagocytosis, but they also activate cytotoxic T lymphocytes (CTLs) to kill infected target cells [1]. T helper 2 cells are a specialized population of T cells that mount a response against extracellular pathogens, such as parasitic worms, that favors the proliferation of CD8 T cells [1]. The Th2 pathway is initiated largely by the presentation of the IL-4 cytokine to a naive Th cell [1], as can be seen in figure 1 below.
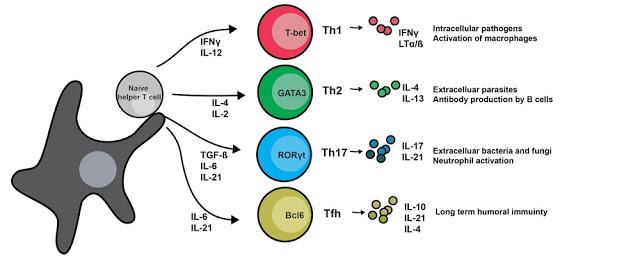
Figure 1. Diagram of T helper cell development via several different pathways. Ligation via different cytokines present in the environment initiates differentiation into several different classes of Th cells [2].
IgD
Unlike IgM but similar to the secretory antibody IgA, IgD has not been shown to recruit pro-inflammatory complement proteins [3]. What IgD is known for thus far is its function as a B cell antigen receptor (BCR), and IgD and IgM receptor engagement by antigen is what induces B cell interactions with T follicular helper cells in the secondary lymphoid tissues [3]. Though its specific role remains elusive, that means that IgD is important in the costimulatory process that leads to the B and T cell proliferation that is so important to the adaptive immune response.
IgD enhances cytokine IL-4 expression in basophils
In order to determine whether IgD levels had an impact on the expression of IL-4 on basophils, the authors used standard and imaging flow cytometry [4] to examine the number of basophils in IgD-deficient mice. IL-4 is a cytokine important in the differentiation of naive T cells in T helper 2 cells [1]. What they found was that IgD-deficient mice had fewer peripheral basophils than the control, and that those basophils expressed less IL-4 [5]. Those IgD-deficient mice also had less IL-4 serum [5], indicating that IgD binds to basophils and increases basophil expression of IL-4.
Using small soluble antigens in the form of food proteins, the researchers showed that they induced plasma cell secretion of IgD. That IgD in turn bound to basophils through the CD44-interacting protein galectin-9, and that ligation of secreted IgD led to the up-regulation of IL-4, IL-5 and IL-13 [5]. Those cytokines are important in the amplification of Th2 cell-mediated responses. CD44 is an antigen expressed in the majority of immune cells and is an important adhesion molecule in the interaction between T lymphocytes and other cells [6].
The next thing Shan et al. discovered was that basophil expression of the cytokine IL-4 increased upon IgD binding to antigen [4]. They demonstrated this by injecting mice that lacked either B cells or both T and B cells with an anti-IgD antibody that induced down-regulation of basophil-bound IgD. Following that injection of the anti-IgD antibody, nonspecific binding of basophil-bound IgD induced expansion and activation of basophils in the spleen, an important secondary lymphoid tissue [4]. The researchers also observed an increase in the expression of IL-4 by those basophils [4]. Additionally, they determined that IgD needed basophils to amplify the humoral Th2 cell-mediated response using qRT-PCR to test for RNA expression of IL-4 in basophil-depleted mice [4].
Having learned this, they then moved on to explore the Th2 cell-amplifying function of IgD. They used an immunization model in which a Th2 cell-inducing and basophil-activating protease (papain) was tested on mice and compared to the response elicited by IgD, and found using flow cytometry and qRT-PCR that mice exhibited comparable basophil requirement in addition to basophil expression of IL-4 [4]. Their findings indicate that IgD employs basophil-activating Th2 cell signals similar to those used by the protease papain to activate basophils [4]. Internalization of papain is a critical component in the polarization of Th2 cells, though there is uncertainty regarding the specific receptors and cell types involved in that papain-induced immune response [7]. At this point, the researchers understood that mouse IgD amplifies papain-induced Th2 cell-mediated responses using a mechanism that involves basophils [4].
Understanding that papain activates basophils in a similar manner to IgD ligand binding by antigen, and knowing that IgD-stimulated basophils increased their expression of the IL-4 cytokine, Shan et al. tested whether papain is capable of activating IgD secretion [4]. They found that papain indeed was able to elicit such a response.
Another important finding from this investigation is that IgD binds to basophil via Galectin-9 and CD44 [4]. This was determined through the use of immunoprecipitation (IP) to study IgD-associated proteins, which were then identified via mass spectrometry. IgD ligation by anti-IgD (as was described earlier) upregulated galectin-9, a degranulation inhibitor and downregulated other degranulation inducers [4]. Shan et al. goes on to discuss how previous studies [8] had demonstrated that galectin-9 is linked to basophils via CD44, thus completing the picture of the IgD binding mechanism important in Th2 cell-mediated response.
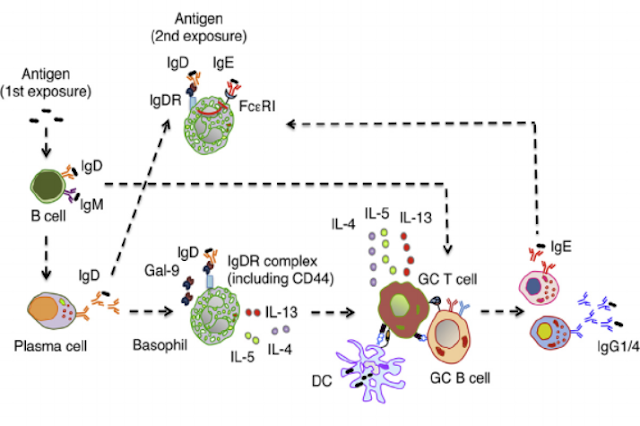
Figure 2. The graphical abstract presented by Shan et al. presents a pictorial depiction of the pathway for IgD in Th2 cell response amplification [4].
Essentially, these findings point to basophils as antigen-presenting cells (APCs) for allergen-induced T helper 2 type response, and illuminate the fact that CD44 and galectin-9 are central components in the Th2 cell response. This presents an exciting new role for IgD, which has long remained the misunderstood class of antibody. These findings have implications for a secondary response to the same antigen, as the IgD Fc receptor complex (IgDR), consisting of CD44 and galectin-9, that was not previously expressed on the basophils before the first antigen encounter will be present during future binding events [4]. Figure 2 above illustrates this concept. In terms of future research and applications for these findings, I would be curious to know whether some sort of injection of IgDR, or at least its major components, would be possible and whether such an approach would speed up the initial Th2 cell-mediated response. Because Th2 cell responses are often associated with allergic reactions and airway inflammation and hyper-reactivity [9], this is very important in potentially finding ways to prevent anaphylaxis in patients exposed to an allergen.
References:
[1] Harrington, L. E., Hatton, R. D., Mangan, P. R., Turner, H., Murphy, T. L., Murphy, K. M., & Weaver, C. T. (2005). Interleukin 17–producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nature immunology, 6(11): 1123-1132.
[2] The Ansel Lab. (2018). MicroRNA regulation of helper T cell differentiation and immune effector functions. University of California San Francisco. https://ansel.ucsf.edu/microrna-regulation-helper-t-cell-differentiation-and-immune-effector-functions
[3] Gutzeit, C., Chen, K., & Cerutti, A. (2018). The enigmatic function of IgD: some answers at last. European journal of immunology. 48: 1101-1113.
[4] Perfetto, S. P., Chattopadhyay, P. K., & Roederer, M. (2004). Seventeen-colour flow cytometry: unravelling the immune system. Nature Reviews Immunology, 4(8): 648-661.
[5] Shan, M., Carrillo, J., Yeste, A., Gutzeit, C., Segura-Garzón, D., Walland, A. C., ... & van de Veen, W. (2018). Secreted IgD Amplifies Humoral T Helper 2 Cell Responses by Binding Basophils via Galectin-9 and CD44. Immunity, 49: 709-724.
[6] Johnson, P., & Ruffell, B. (2009). CD44 and its role in inflammation and inflammatory diseases. Inflammation & Allergy-Drug Targets (Formerly Current Drug Targets-Inflammation & Allergy), 8(3), 208-220.
[7] Dwyer, D. F., Woodruff, M. C., Carroll, M. C., Austen, K. F., & Gurish, M. F. (2014). B Cells Regulate CD4+ T Cell Responses to Papain following B Cell Receptor–Independent Papain Uptake. The Journal of Immunology, 1303247.
[8] Niki, T., Tsutsui, S., Hirose, S., Aradono, S., Sugimoto, Y., Takeshita, K., ... & Hirashima, M. (2009). Galectin-9 is a high affinity IgE-binding lectin with anti-allergic effect by blocking IgE-antigen complex formation. Journal of Biological Chemistry, jbc-M109.
[9] Hansen, G., Berry, G., DeKruyff, R. H., & Umetsu, D. T. (1999). Allergen-specific Th1 cells fail to counterbalance Th2 cell–induced airway hyperreactivity but cause severe airway inflammation. The Journal of clinical investigation, 103(2), 175-183.
No comments:
Post a Comment